

Cours de SVTHEEB Terminale D – Les mécanismes fondamentaux de la reproduction sexuée chez les mammifères et les spermaphytes : La fécondation
III. Gamètes et fécondation
Chez tous les êtres vivants qui se reproduisent par voie sexuée, la fécondation est articulée autour des étapes suivantes :
– Rapprochement des gamètes ;
– Pénétration du gamète mâle ;
– Caryogamie ou fusion des pronoyaux mâles et femelle (fécondation).
III.1- Rapprochement des gamètes.
Pour que la fécondation ait lieu, il faut que les gamètes mâles et femelles se rencontrent.
Chez les Mammifères, cette rencontre est facilitée par :
– le rapprochement des sexes ;
– l’accouplement ;
– la mobilité du spermatozoïde ;
– les contractions de la paroi et la vibration des cils tapissant la paroi des voies génitales féminines.
Chez les Spermaphytes, la rencontre entre le gamète mâle et le gamète femelle nécessite une pollinisation c’est-à-dire un transfert de pollen de l’étamine d’une fleur au stigmate du pistil (chez les angiospermes) ou à l’ovule (chez les gymnospermes) de la même fleur (autogamie) ou d’une fleur différente (allogamie).
Les agents de la pollinisation sont variés (vent, eau, êtres vivants). La pollinisation est suivie du développement du tube pollinique.
III.2- Pénétration du gamète mâle.
III.2.1. Chez les Mammifères.
La pénétration du gamète mâle, prélude à la fusion des noyaux, se fait suivant le mécanisme ci-après :
Chronologie des évènements
• L’ovocyte II est bloqué en métaphase II.
• Plusieurs spermatozoïdes s’infiltrent entre les cellules folliculaires et viennent au contact de la zone pellucide.
• Certaines protéines de la zone pellucide fixent sélectivement les protéines complémentaires de la membrane de la tête du spermatozoïde.
• Cette liaison en déstabilisant la membrane et l’acrosome des spermatozoïdes, permet la libération d’enzymes digérant la zone pellucide, facilitant ainsi leur progression vers la membrane du gamète femelle.
• Arrivé au contact de la membrane de l’ovocyte, un spermatozoïde va être phagocyté par l’ovocyte. En même temps, les granules corticaux déchargent par exocytose leur contenu enzymatique, ce qui provoque un durcissement de la zone pellucide et un masquage des protéines de fixation des spermatozoïdes. Cette réorganisation évite la polyspermie.
• Les cellules folliculaires se sont rétractées, participant ainsi au blocage de la polyspermie.
• Rétraction du cytoplasme ovocytaire et réveil physiologique de l’ovocyte : reprise des synthèses, respiration, méiose…
• L’espace péri-ovocytaire apparaît nettement.
• Formation du second globule polaire.
• Le noyau du spermatozoïde gonfle et devient le pronucléus mâle. Le reste du gamète est digéré par l’ovocyte, sauf le centriole proximal qui formera l’aster.
• Formation du pronucléus femelle et migration des deux pronucléi l’un vers l’autre.
• Fusion des pronucléi en prophase : c’est la caryogamie.
• Formation de l’œuf ou zygote à 2n chromosomes.
• Anaphase de la première mitose de l’œuf.
III.2.1.1 Evolution du taux d’ADN au cours de la fécondation
L’évolution de la masse totale d’ADN que contient l’ovocyte II au cours de la fécondation permet de construire le graphe suivant :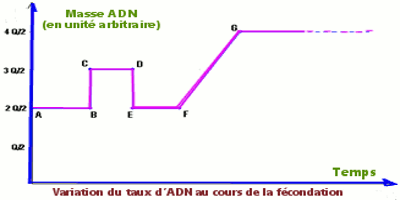
– Le segment AB représente la phase où l’on a un ovocyte II à n chromosomes à deux chromatides chacun.
– Le segment BC traduit l’entrée d’une masse Q/2 d’ADN, ce qui équivaut à l’entrée brusque (l’irruption) d’un spermatozoïde avec n chromosomes à une chromatide chacun.
– Le segment CD correspond au rapprochement des pronucléi et à l’activation de l’ovocyte II.
– Le segment DE traduit la perte d’une masse Q/2 d’ADN correspondant à un jeu de n chromosomes à une chromatide chacun. Ceci ne peut être dû qu’à l’expulsion du second globule polaire.
– Le segment EF correspond à une masse Q d’ADN, ce qui est dû aux n chromosomes non dupliqués apportés par l’ovotide.
– Le segment FG traduit la duplication de la masse d’ADN, ce qui prépare la première mitose de la cellule œuf.
III.2.2- Chez les Spermaphytes.
Le noyau reproducteur subit une mitose et engendre deux gamètes mâles allongés et plus ou moins spiralés : les anthérozoïdes. Le noyau végétatif, après avoir dirigé la croissance du tube pollinique disparaît au voisinage de l’ovule.
Quand les deux anthérozoïdes pénètrent dans le sac embryonnaire, l’un féconde l’oosphère engendrant ainsi un œuf diploïde appelé œuf principal ou œuf embryon. L’autre anthérozoïde s’unit aux deux noyaux du sac et engendre un œuf triploïde appelé œuf accessoire. Cette double fécondation est caractéristique des Angiospermes (Spermaphytes à graine enveloppée).
Par la suite, les synergides et les cellules antipodes dégénèrent. Il ne subsiste dans le sac que les deux œufs. L’ovule se transforme ainsi en graine et l’ovaire en fruit.
Conclusion
La reproduction sexuée est marquée par l’alternance régulière de deux mécanismes biologiques :
– la méiose qui assure le passage de l’état diploïde à l’état haploïde ;
– la fécondation qui réunit les deux lots haploïdes des chromosomes des gamètes pour rétablir l’état diploïde.
Le bagage chromosomique caractéristique de l’espèce est ainsi conservé génération après génération. Le même caryotype, diploïde par exemple, est reconstitué à chaque nouvelle fécondation. On parle de cycle chromosomique pour évoquer cette reconstitution périodique du stock chromosomique caractéristique de l’espèce.
Les positions relatives de la méiose et de la fécondation varient selon les espèces :
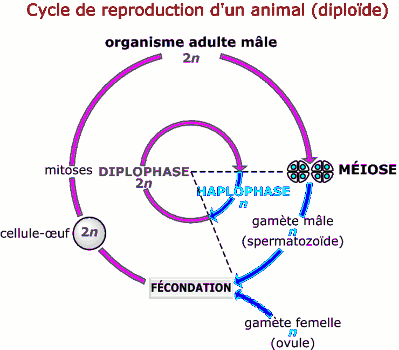
• Chez les animaux et certains végétaux (algues brunes par exemple), la méiose intervient au moment de la gamétogenèse. Les cellules sexuelles sont les seules cellules haploïdes. Après la fécondation, les cellules du nouvel individu qui descendent de la cellule œuf par mitoses sont diploïdes (2n). Un tel cycle de développement où la méiose précède la fécondation est caractérisé par la prédominance remarquable de la phase diploïde : il est dit diplophasique.
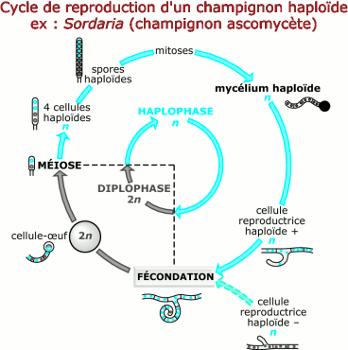
• Chez certains végétaux inférieurs (Champignon Neurospora ou Sordaria par exemple), la méiose succède immédiatement à la fécondation. La phase diploïde dans ce cas est réduite à la cellule œuf. Dans un tel cycle, les individus sont constitués de cellules à n chromosomes ; c’est la phase haploïde qui prédomine : c’est un cycle haplophasique.
• Chez de nombreux végétaux (mousses, fougères, angiospermes, …), méiose et fécondation sont nettement séparés au cours du cycle de développement. La cellule œuf se développe en un ensemble de cellules diploïdes. Au sein de cet ensemble, certaines cellules subissent la méiose et donne naissance à des cellules haploïdes qui se développent en un organisme haploïde responsable de la production des gamètes. On observe donc l’alternance d’une phase haploïde et d’une phase diploïde ; l’importance de chacune variant d’une espèce à l’autre.
Chez certains êtres, pour passer d’une génération à l’autre se succèdent des cellules haploïdes et des cellules diploïdes formant l’haplophase et la diplophase. Conventionnellement, on représente ces deux phases successives comme traçant un cycle perpétuel : le cycle chromosomique.
On appelle cycle de reproduction ou cycle biologique d’une espèce, la succession des étapes suivies par les organismes depuis la formation de l’œuf jusqu’à l’état adulte et la formation d’une nouvelle génération à partir des gamètes parentaux.
La transition entre diplophase et haplophase se fait à travers la méiose; tandis que la transition entre haplophase et diplophase se traduit à travers la caryogamie ou fusion des noyaux à l’issue de la fécondation.
Selon tes organismes, l’importance est relative entre les deux phases.
Chez les animaux, le diplophase est prédominante (la méiose précède la fécondation et on parle de cycle diplophasique ou diplobiontique); tandis que de nombreux champignons (ascomycètes) ont un cycle à haplophase prédominante (la méiose suit immédiatement la fécondation et le cycle est dit haplophasique ou haplobiontique).



Chez le Polypode la méiose intervient lors de la formation des spores; la spore est donc une cellule haploïde.
Le prothalle, né de la spore est donc lui aussi haploïde; il produit les gamètes et est le gamétophyte.
La Fougère, née de la fécondation est par conséquent diploïde ; elle produit des spores et constitue le sporophyte.
Le cycle de cette plante est marqué par la succession de deux êtres (le sporophyte et le gamétophyte} coïncidant avec la succession de deux phases chromosomiques (la diplophase et l’haplophase) : c‘est un cycle haplodiplophasique.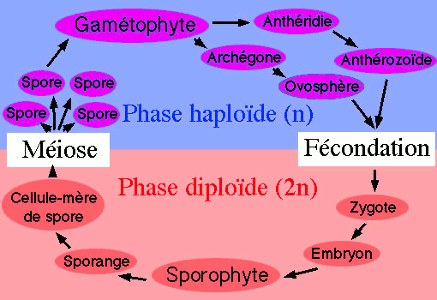
Titre : Cycle de développement des spermaphytes (Cycle haplodiplophasique)
Chez les spermaphytes, par comparaison au cycle de la fougère, on retrouve la succession de deux générations :
– Une génération diploïde qui se termine par la formation des micro et macrospores et qui représente le sporophyte (c’est la plate feuillée et fleurie).
– Une génération haploïde (qui se termine par l’union des gamètes et qui représente le gamétophyte (prothalle).
Ce cycle est donc haplodiplophasique ou haplodiptobiontique.
On note cependant quelques différences :
– Les organes produisant les gamètes sont protégés dans une fleur,
– Il y a deux catégories de prothalle unisexué et minuscule (le prothalle mâle est le grain de pollen et le prothalle femelle est le sac embryonnaire qui reste inclus dans le sporophyte sur lequel il vit en parasite
– La fécondation est double et indépendante de l’élément liquide,
– Enfin les œufs se développent à l’intérieur de l’ovule, donc aux dépens du sporophyte.
Ainsi chez les végétaux, la reproduction sexuée a des caractères fondamentalement semblables à celle des animaux.
La méiose et la fécondation permettent le maintien de la garniture chromosomique (à la fois le nombre et les caractéristiques des chromosomes) de l’espèce et donc sa stabilité.

